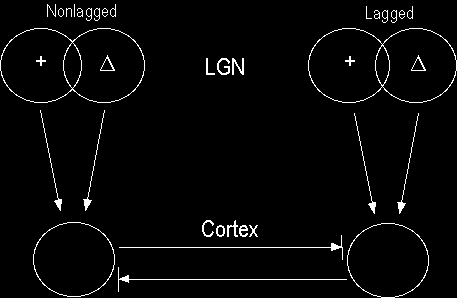
Two mutually inhibitory simple cells, each of which receives ON and OFF inputs from the LGN. If these inputs are in spatiotemporal quadrature, the simple cells will form a push-pull pair (in the preferred direction). The figures below show what the responses to sinusoidally-modulated bars would look like in this case.
The LGN inputs (in black and white) are spatiotemporally separable. In this particular example, I chose to have nonlagged ON and OFF cells project to one cortical cell and lagged ON and OFF inputs project to the other cortical cell. Other combinations are equally acceptable, but this choice illustrates the points I wanted to make clearly. The cortical cells here receive inputs that are in spatiotemporal quadrature.
The cortical responses are computed from the excitation provided by the LGN inputs, with the inhibitory input from the other cortical cell subtracted. Because there is feedback in this anatomy, the calculations are made by relaxing the system until it converges.
Even though each cortical cell receives separable inputs from the LGN, the relayed quadrature inputs make them direction selective. The two mutually inhibitory cortical cells share the same spatiotemporal orientation, so that they prefer the same direction of motion. The take-home message here is that cells that inhibit each other have similar response properties.
The cortical cells are in a push-pull arrangement for the preferred direction, firing a half-cycle out of phase with each other. In the nonpreferred direction, the cells fire at the same time, thereby suppressing each other's responses. From the point of view of the space-time maps, the receptive fields are not really in a push-pull configuration in the sense that the responses occur a half-cycle apart, but instead they are a quarter-cycle apart in space and in time. The receptive fields are complementary, which inseparable fields can be only if they share a common orientation.
I assume here that the effect of adapting is to potentiate inhibition. Thus, to simulate the adapted state I simply increased the inhibitory gain between the cortical cells. This assumption has not been supported by evidence. One current line of thought (cf. Findlayson and Cynader, ref to come) is that presynaptic metabotropic glutatmate receptors are involved in synaptic depression that underlies adaptation. The present results provide a challenge for theoretical accounts, and I look forward to seeing an improved model that is built on empirically supported mechanisms. For now, however, this model is at least consistent with the data.
There is an additional assumption here, which is that the inhibition is low-pass filtered relative to the firing of the cell that provides it. This reasonable assumption permits the onset of the response to be affected, but the offset is untouched because the source of the inhibition is not active just prior to offset.

Figure 11
Figure 12 shows the model's predictions for the receptive field maps, to compare to the actual data in Figure 7. Only certain positions are affected.

Figure 12
Figure 13 presents the model response to a grating drifting in the preferred direction. As in the actual data of figures 1 and 6, only onset is affected.

Figure 13
Next section is Conclusions.
Back to the Index.